Нервово-м'язові веретена є рецепторами розтягування, розташованими в межах добровільної мускулатури; з їхньою діяльністю вони здатні захопити стан розтягування м'язів і направити інформацію, зібрану до спинного мозку і до енцефалону. Діяльність нервово-м'язових веретен є дуже важливою для запобігання травм, пов'язаних з надмірним подовженням, як для підтримки нормального м'язового тонусу, так і для гармонійного та контрольованого руху рідини.
Всі скелетні м'язи, за винятком нижньощелепної м'язи, містять у собі різні нервово-м'язові веретена, які особливо концентруються на рівні жувальних м'язів, хребта, очей, кінцівок і рук. Тут нервово-м'язові веретени довжиною близько 5-10 мм розташовуються паралельно з звичайними м'язовими волокнами, і завдяки цій «побічній» компоновці вони здатні захоплювати ступінь подовження.
анатомія
Нервово-м'язовий веретен складається з капсули сполучної тканини, яка обертається навколо невеликої групи м'язових волокон (від 4 до 10), оснащених «особливою» цитологічною структурою; ці волокна часто називають интрафузными, щоб відрізнити їх від звичайних, які, для рівних ігрових сфер, дають прикметник "екстрафусалі".
Фізіологія внутрішньофазних волокон пояснюється, насамперед, детально вивченням анатомічної структури. На їхніх крайностях вони досить схожі на звичайні волокна і для цього містять скоротливі смугасті фібрили. Реальна відмінність полягає в екваторіальній частині, яка виглядає збільшеною, позбавленою міофібрил і багатою чутливими до розтягунь сенсорними закінченнями, зануреними в желатинову речовину.
Сказано, отже, що волокна нервово-м'язових веретенів є ефекторними до двох полюсів (вони скорочуються у відповідь на нервовий стимул) і випромінювачами в центрі (з яких вони посилають інформацію про стан подовження).
З анатомічної точки зору інтрафузійні м'язові волокна поділяються на ядерні волокна мішка (також називаються волокна мішка або мішка) і волокна ядерного ланцюга. Перші мають розширену центральну площу, багату ядрами. З іншого боку, волокна ядерного ланцюга мають подовжене ядерне розподіл, завжди зосереджене в екваторіальній області, але також розширені в периферії; вони також коротше і тонше, ніж попередні.
З анатомічної точки зору чутливі закінчення нервово-м'язового веретена влаштовані, частково шляхом згортання до серединної області (anulum-спіраль або первинні закінчення) і частково формування гілки розсади у сусідніх областях (квіткові або вторинні закінчення).
Первинні закінчення товщі, мають високу швидкість провідності, належать до класу волокон Ia і відходять від волокон мішка і ядерного ланцюга; вторинні закінчення, що належать до класу волокон типу II, замість тонше, менш швидкі при поширенні імпульсів і іннервують переважно ланцюгові волокна ядер.
З фізіологічної точки зору, з іншого боку, ми виділяємо швидкодіючі чутливі волокна (тип Ia) і чутливі до повільного провідності волокна (тип II). Перші, незважаючи на припинення обох типів волокон, є анулярно-спіральними закінченнями, характерними для волокон мішечного волокна динамічних ядер (див. Нижче). З іншого боку, більш повільні волокна II мають анулярно-спіральні закінчення, які обертають волокно мішка-волокно статичних ядер і волокон ланцюга; до цієї категорії відносяться також цвітіння.
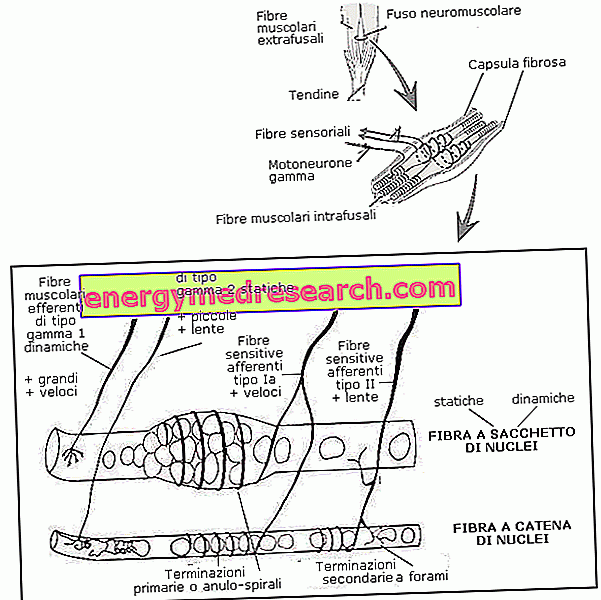
На відміну від екстрафузальних м'язових волокон, які отримують афферентні лінії від альфа-моторних нейронів, шпиндельні волокна піддаються дії гамма-мотонейронів (нервові волокна, що надходять з переднього рога спинного мозку, характеризуються малим калібром).
ПРОДОВЖИТИ: Фізіологія нервово-м'язових веретен »