Щоб мати можливість всебічно говорити про гемоглобін (Hb), корисно спершу мати справу з міоглобіном (Mb), який дуже схожий на гемоглобін, але набагато простіше. Існують жорсткі родинні зв'язки між гемоглобіном і міоглобіном: обидва є кон'югованими білками і їх простетична група (небілкова частина) є групою гема .
Міоглобін є глобулярним білком, що складається з єдиного ланцюга з приблизно ста п'ятдесяти амінокислот (залежно від організму), і його молекулярна маса становить близько 18 Kd.
Як згадувалося, він обладнаний гем-групою, яка вставлена в гідрофобну (або ліпофільну) частину білка, що складається з фальцювань, що відносяться до α-спіральних структур волокнистих білків.
Міоглобін складається в основному з сегментів α-спіралей, присутніх у кількості восьми і складається, майже виключно, з неполярних залишків (лейцин, валін, метіонін і фенілаланін), а полярні залишки практично відсутні (аспарагінова кислота, глутамінова кислота, лізин і аргінін); єдиними полярними залишками є два гістидину, які відіграють фундаментальну роль у атаці кисню до групи гема.
Гем-група є хромофорною групою (поглинається у видимій) і є функціональною групою міоглобіну.
Див. Також: глікозильований гемоглобін - гемоглобін у сечі
Трохи хімії
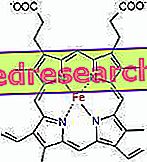
Зв'язок між протопорфирином і залізом є типовим зв'язком так званих координаційних сполук, які є хімічними сполуками, в яких центральний атом (або іон) утворює зв'язки з іншими хімічними видами в кількості, що перевищує його число окислення (електричний заряд). У випадку гема ці зв'язки є оборотними і слабкими.
Координаційне число (кількість координаційних зв'язків) заліза - шість: навколо заліза може бути шість молекул, які поділяють електронні зв'язки.
Щоб сформувати координаційну сполуку, вона приймає дві орбіталі з правильною орієнтацією: людина може "купувати" електрони, а інша здатна їх пожертвувати
У гемі залізо утворює чотири плоских зв'язки з чотирма атомами азоту, які знаходяться в центрі прото-порфиринового кільця, і п'ятий зв'язок з проксимальним гістидиновим азотом; залізо має шосту вільну координаційну зв'язок і може зв'язуватися з киснем.
Коли залізо знаходиться у формі вільного іона, його орбіталі d- типу мають однакову енергію; в міоглобіні іон заліза пов'язаний з протопорфирином і гістидином: ці види магнітно порушують орбіталі заліза; ступінь збурень буде різною для різних орбіталей відповідно до їх просторової орієнтації та загрозливих видів. Оскільки сумарна енергія орбіталей повинна повертатися до постійної, збурення викликає енергетичне поділ між різними орбіталями: енергія, отримана деякими орбіталями, еквівалентна енергії, втраченої іншими.
Якщо поділ, що відбувається між орбіталями, не дуже великий, краще використовувати електронну систему з високим спіном: електронні зв'язки намагаються організувати їх паралельно у декількох можливих під-рівнях (максимальна кратність); якщо, з іншого боку, збурення дуже сильне і існує велике розмежування між орбіталями, то може бути зручніше поєднувати зв'язуючі електрони в нижніх енергетичних орбіталях (низький спін).
Коли залізо зв'язується з киснем, молекула набуває низький рівень спина, тоді як залізо має шосту безкоштовну координаційну ланку, молекула має високий спин.
Завдяки цій різниці в спині, через спектральний аналіз міоглобіну, ми можемо зрозуміти, чи є пов'язаний кисень (MbO2) чи ні (Mb).
Міоглобін є білком, типовим для м'язів (але не тільки в м'язах).
Міоглобін витягується з кашалота, в якому він присутній у великих кількостях і потім очищається.
Китоподібні мають дихання, як у людей: маючи легені, вони повинні поглинати повітря через дихальний процес; кашалот повинен містити якомога більше кисню в м'язах, які здатні накопичувати кисень, зв'язуючи його з міоглобіном; Кисень потім повільно вивільняється, коли китоподібних занурюється, тому що його метаболізм вимагає кисню: тим більше кількість кисню, що кашалот здатний поглинати, і більше кисню доступно під час занурення.

Зробити м'ясо більш-менш червоним - це вміст гемопротеїнів (саме гем робить червоне м'ясо).
Гемоглобін має багато структурних аналогій з міоглобіном і здатний оборотно зв'язувати молекулярний кисень; але, хоча міоглобін приурочений до м'язів і периферичних тканин в цілому, гемоглобін міститься в еритроцитах або еритроцитах (вони є псевдо-клітинами, тобто вони не є справжніми клітинами), які складають 40% крові.
Гемоглобін є тетраметровим, тобто складається з чотирьох поліпептидних ланцюгів, кожна з гемовою групою і два на два (у людини є дві альфа-ланцюги і дві бета-ланцюги).
Основною функцією гемоглобіну є транспорт кисню ; ще однією функцією крові, в якій бере участь гемоглобін, є транспортування речовин до тканин.
На шляху від легких (багатих киснем) до тканин гемоглобін переносить кисень (в той же час інші речовини також потрапляють до тканин), у той час як на зворотному шляху він несе в собі відходи, зібрані з тканин, особливо діоксид вуглецю, що утворюється в метаболізмі.
У розвитку людини існують гени, які виражаються лише протягом певного періоду часу; з цієї причини у нас є різні гемоглобіни: ембріональний, ембріональний, дорослої людини.
Ланцюги, що складають ці різні гемоглобіни, мають різні структури, але з подібністю функція, яку вони виконують, більш-менш однакова.
Пояснення присутності декількох різних ланцюгів відбувається наступним чином: під час еволюційного процесу організмів гемоглобін також розвинувся, спеціалізуючись на транспортуванні кисню з районів, багатих дефіцитними районами. На початку еволюційного ланцюга гемоглобін переносив кисень у дрібні організми; в ході еволюції організми досягли більших розмірів, тому гемоглобін змінився, щоб мати можливість транспортувати кисень до районів, розташованих далі від того, що він був багатий; Для цього в ході еволюційного процесу були кодифіковані нові структури ланцюгів, що становлять гемоглобін.
Міоглобін зв'язує кисень навіть при скромному тиску; в периферичних тканинах спостерігається тиск (РО2) близько 30 мм рт.ст.: при цьому тиску міоглобін не вивільняє кисень, тому він неефективний як кисневий носій. Гемоглобін, з іншого боку, має більш еластичну поведінку: він зв'язує кисень при високих тисках і вивільняє його, коли тиск знижується.
Коли білок функціонально активний, він може трохи змінити свою форму; наприклад, міоглобін з оксигеном має іншу форму від міоглобіну без кисню, і ця мутація не впливає на сусідні.
Дискурс відрізняється у випадку асоційованих білків, таких як гемоглобін: коли ланцюг окислюється киснем, він індукується для зміни форми, але ця модифікація є тривимірною, тому інші ланцюги тетраметра також впливають. Той факт, що ланцюги пов'язані один з одним, наводить на думку, що модифікація одного впливає на інших сусідніх, навіть якщо в різній мірі; коли ланцюг окислюється киснем, інші ланцюги тетраметра приймають «менш вороже ставлення» до кисню: труднощі, з якими ланцюг оксигенатів зменшується, коли ланцюги поруч з ним киснюються. Той самий аргумент справедливий для дезоксигенації.
Четвертинна структура дезоксигемоглобіну називається T (напруженою) формою, а оксигемоглобіну - R- формою (вивільняється); в розтягнутому стані існує ряд досить сильних електростатичних взаємодій між кислотними амінокислотами і основними амінокислотами, які призводять до жорсткої структури дезоксигемоглобіну (тому "напружена форма"), тоді як, коли кисень пов'язаний, сутність цих взаємодії зменшуються (тому "випущена форма"). Крім того, за відсутності кисню заряд гістидину (див. Структуру) стабілізується протилежним зарядом аспарагінової кислоти, в той час як в присутності кисню спостерігається тенденція до втрати білка протона; все це означає, що оксигенований гемоглобін є сильнішою кислотою, ніж деоксигенована гемоглобія: ефект бору .
Залежно від рН, гем-група більш-менш легко зв'язується з киснем: в кислому середовищі гемоглобін легше вивільняє кисень (напружена форма стабільна), а в основній середовищі зв'язок з киснем - сильніше.

Кожен гемоглобін випускає 0, 7 протонів на вхідний моль кисню (O2).
Ефект Бора дозволяє гемоглобіну поліпшити його здатність переносити кисень.
Гемоглобін, що робить подорож від легенів до тканин, повинен врівноважувати себе відповідно до тиску, рН і температури.
Ми бачимо вплив температури .
Температура в легеневих альвеолах на 1-1, 5 ° С нижче зовнішньої температури, а в м'язах температура становить 36, 5-37 ° С; з підвищенням температури коефіцієнт насичення зменшується (при тому ж тиску): це відбувається тому, що кінетична енергія зростає, і дисоціація є сприятливою.

Існують і інші фактори, які можуть впливати на здатність гемоглобіну зв'язуватися з киснем, одним з яких є концентрація 2, 3 бісфосфоглицерата .
2, 3-бісфосфогліцерат є метаболічним агентом, що присутній в еритроцитах в концентрації 4-5 мМ (в жодній іншій частині тіла він не присутній в такій високій концентрації).
При фізіологічному рН 2, 3 бісфосфогліцерат депротонируется і має на ньому п'ять негативних зарядів; вона вклинюється між двома бета-ланцюгами гемоглобіну, оскільки ці ланцюги мають високу концентрацію позитивних зарядів. Електростатичні взаємодії між бета-ланцюгами і бісфосфогліцератними ланцюгами надають системі певну жорсткість: отримана напружена структура, яка має невелику спорідненість до кисню; під час оксигенації, потім, 2, 3 бісфосфогліцерат витісняється.
У еритроцитах є спеціальний апарат, який перетворює 1, 3-бісфосфогліцерат (утворюється в результаті метаболізму) в 2, 3-бісфосфогліцерат таким чином, що він досягає концентрації 4-5 мМ і тому гемоглобін здатний обмінюватися кисню в тканинах.
Гемоглобін, що надходить у тканину, знаходиться у вивільненому стані (зв'язаний з киснем), але в околиці тканини він карбоксильований і переходить у розтягнуте стан: білок в цьому стані має більш низьку тенденцію до зв'язування з киснем, порівняно з вивільненим станом, тому гемоглобін вивільняє кисень до тканини; крім того, внаслідок реакції між водою і діоксидом вуглецю відбувається виробництво іонів H +, отже, додатковий кисень обумовлений ефектом бору.
Діоксид вуглецю дифундує в еритроцит, що проходить через плазматичну мембрану; Враховуючи, що еритроцити становлять близько 40% крові, слід очікувати, що тільки 40% діоксиду вуглецю, який дифундує з тканин, потрапляє в них, насправді 90% діоксиду вуглецю надходить у червоні клітини, оскільки вони містять фермент, який перетворює двоокису вуглецю в вуглекислоті випливає, що стаціонарна концентрація вуглекислого газу в еритроцитах низька і тому швидкість входу висока.

Інше явище, яке відбувається, коли еритроцит досягає тканини, наступне: за градієнтом, HCO3- (похідне діоксиду вуглецю) виходить з еритроцита і, щоб збалансувати вихід негативного заряду, ми маємо вхід хлоридів, що визначає збільшення осмотичного тиску: щоб врівноважити цю зміну, виникає також і надходження води, що викликає набряк еритроцитів (ефект HAMBURGER). Протилежне явище виникає, коли еритроцит досягає легеневих альвеол: відбувається дефляція еритроцитів (ефект HALDANE). Таким чином, венозні еритроцити (безпосередньо до легенів) є округлими, ніж артеріальні.


